
近日,复旦大学智能复杂体系基础理论与关键技术实验室于玉国教授课题组探讨了人脑在执行心算和聆听音乐两种认知行为时呈现出来的动态脑网络复杂动力学特征,揭示了人脑在执行高度专注的认知任务时多个脑区形成稳健的动态长程关联网络特征,而在听舒缓音乐时大脑则呈现出局部中心化的特征连接模式。该研究成果已在线发表于《表型组学》。
人脑在放松的静息态或听音乐状态和专注工作的任务态下可能有着截然不同的信息交互模式。大脑通常会调动多个脑区协同配合才能完成一些高度复杂的认知任务,在这种任务状态时,大脑多个脑区形成的功能网络具有什么样的特征?网络构型随时间变化存在什么样的规律?与此对应,人在听音乐的放松状态下,大脑网络又会表现出什么样的结构和信息传输规律?在这些有趣问题的驱使下,研究人员展开了这项研究。
多通道脑电记录技术因其具有无创性,高时间分辨率和直接的神经活动反映等特点,在认知神经科学和脑机接口等领域具有广泛的应用。该文研究者设计了脑电认知实验,应用64通道脑电对人脑在闭眼静息、睁眼注视、听音乐和做心算任务时的脑电活动分别进行系统记录,并应用图论(如最小生成树分析方法,见图1)分析了脑电的时空动力学特征。
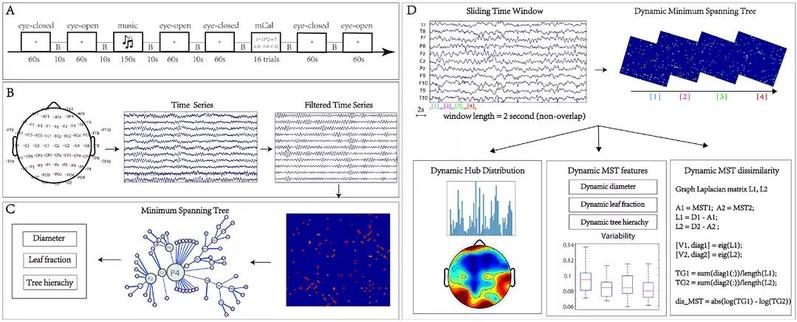
图1 | 实验范式和数据分析流程图。(A) 该文采用的脑电实验范式;(B) 脑电预处理过程;(C) 静态最小生成树分析流程,包括计算树直径、叶子分数和树层级这三个生成树特征;(D) 动态最小生成树分析过程,作者采用2s非重叠的滑动时间窗方法构建动态生成树网络,计算了随时间变化的动态核心节点分布图,动态最小生成树特征变异性和动态最小生成树不相似性指数。
以往的脑电网络研究大多关注静态的特征改变,很少揭示其动态的规律。为此,研究人员基于不依赖阈值的最小生成树分析方法,构建动态脑网络。为了研究在不同状态下是否存在始终扮演着重要角色的核心脑区,研究人员统计了随时间动态变化过程中,每层网络排名前五的核心节点(hub)的出现频次,将其投射到脑地形图上,直观呈现出随时间变化过程中哪些脑区始终扮演着重要的角色。此外,研究者还计算了基于图拉普拉斯变换的动态不相似性指数,刻画最小生成树网络随时间演变的拓扑结构变化规律。
在平均网络连通度为4的拓扑连接情况(图2A),闭眼状态下,脑功能连接主要集中在后顶枕区,右侧连接比左侧更紧密。在听音乐状态下,连接也分布在后顶枕区,与中央区有少量连接,左右脑分布相对均匀。在睁眼状态下,连接变得随机并呈现全脑弥散。心算状态下,脑连接双侧对称分布,加强了左右脑(尤其是双侧额叶、颞叶和顶叶)之间的信息交流。
权重网络分析结果显示(图2B,C),在全频段、alpha频段和beta频段,闭眼静息、睁眼静息、听音乐和做心算四种状态下,脑网络特征(功能连接、聚类系数、特征路径长度)存在显著性差异。其中,听音乐时的脑功能连接最强,而心算时功能连接较弱。听音乐时网络聚类系数最大,特征路径长度最小,而心算时聚类系数较小,特征路径较大。
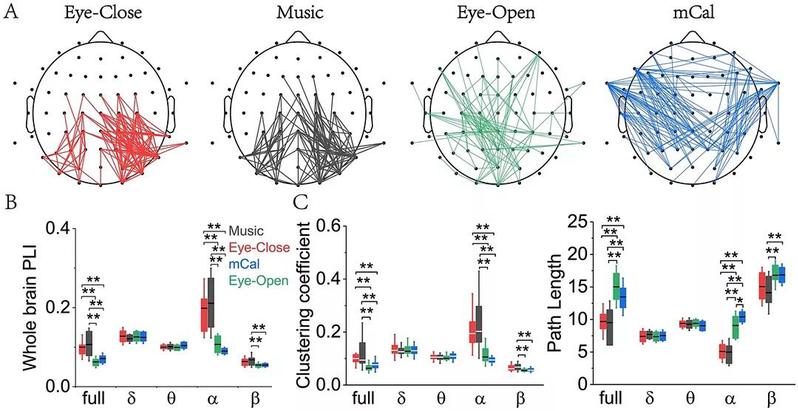
图2 | 权重网络分析相关参数。(A) 网络平均度为 4 时的功能连接模式;(B) 不同频段的全脑功能连接 (PLI) 对比;(C) 不同频段的聚类系数和特征路径长度。alpha显著性水平设置为0.05,当p小于0.05 (但大于0.01)时用单星号标识,当p小于0.01时用双星号标识。
应用图论中最小生成树分析方法得到的结果显示(图3),在全频段、alpha频段,闭眼静息、睁眼静息、听音乐和做心算四种状态下的三个生成树特征(树层级tree hierarchy, TH,叶子分数leaf fraction, LF,和树直径 diameter, D)存在显著性差异。树层级度量了网络节点拥有多个子功能网络的特征;树直径度量了网络的最长路径长度,树直径越大表明网络中存在越长的距离传输;叶子分数度量了网络中度为1的节点占总节点数的比例。叶子分数越大表明网络趋于中心化的星型结构,叶子分数越小表明网络趋于线型结构。分析发现,听音乐时叶子分数最大,树层级大,树直径最小,网络连接趋于星型树结构,而心算时叶子分数最小,树层级小,树直径较大,网络连接趋于线型结构。
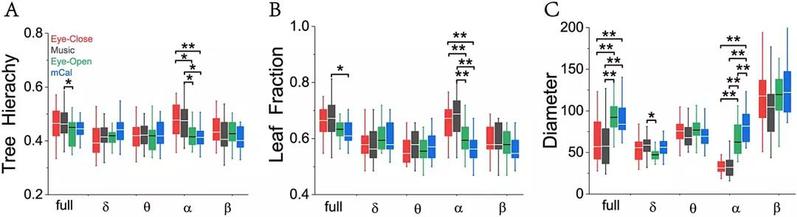
图3 | 不同频段最小生成树统计特征:(A) 树层级 (TH);(B) 叶子分数;(C) 树直径。
考虑四种状态的网络特征在alpha频段存在显著性差异,该文绘制了该频段下的最小生成树连接模式图(图4),以更好地呈现最小生成树的拓扑结构。从图4A中,很容易发现四种状态下最小生成树的核心节点截然不同,其中,在闭眼状态下,CP2, P3, P4, C2, CP4, P6, PO7等节点扮演着比较重要的角色;而听音乐时P4, P8, O2, P2等节点扮演着重要的角色。对比之下,在睁眼状态下,FC2, CP1, CP2, FCz, FC4等节点扮演着重要的角色,而心算时CP1, Pz, AFz, FCz, C2扮演着重要的角色。图4B显示,听音乐时的连接模式趋向于星型结构,而睁眼和心算时的连接模式趋于线型结构;尤其是心算状态,所有的核心节点都集中在网络骨干线上,相较于睁眼状态更趋于线型结构。
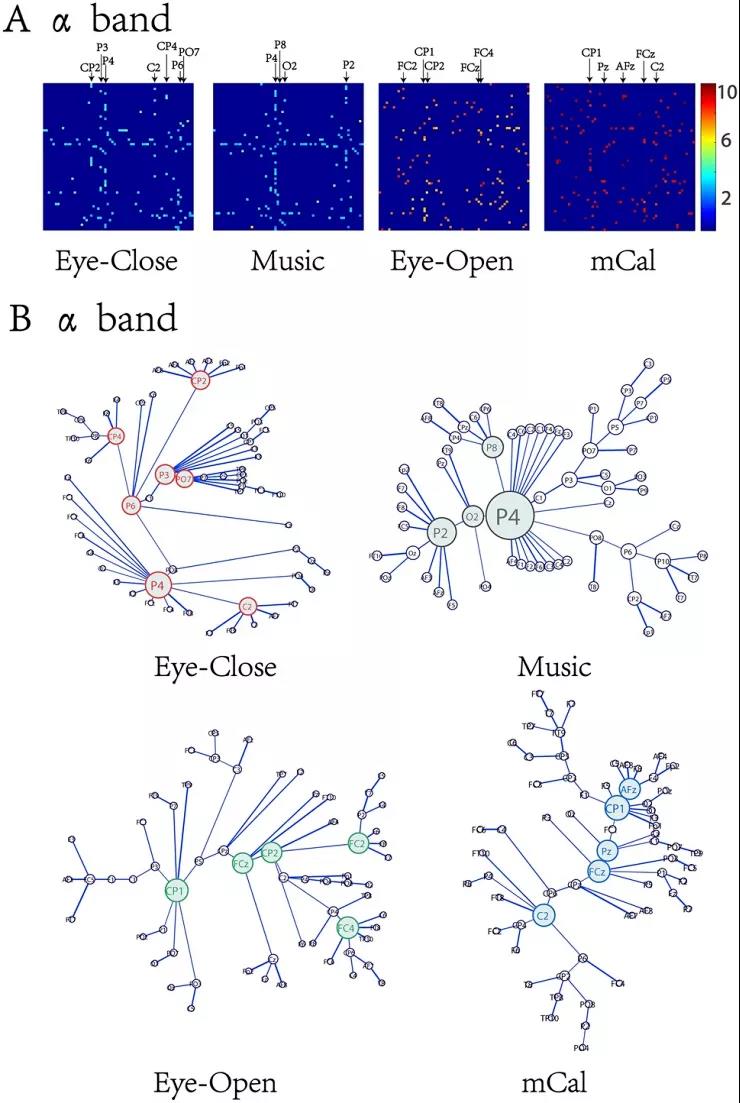
图4 | α频段中四种状态下的最小生成树拓扑连接图。(A) 最小生成树连接矩阵图,图上标识的电极为核心节点;(B) 最小生成树拓扑连接图。有颜色的节点表示核心节点,更大的节点意味着在网络信息传输中扮演着更重要的角色。
所有被试的动态最小生成树核心节点分布图显示(图5),存在一些关键的脑区,随时间演变始终扮演着重要的角色。结合最小生成树中随时间变化过程网络连接度最大的5个节点分布图和频率地形图,可以很容易看到,不同行为状态下始终扮演关键角色的脑区有所不同。在闭眼和听音乐的状态下,少数前额叶、双侧颞叶和大部分双侧后头部区域(包括顶叶和枕叶)更频繁地起到核心节点作用。然而,在睁眼和心算状态下,最小生成树的核心节点经常出现在前额叶和双侧颞顶叶。
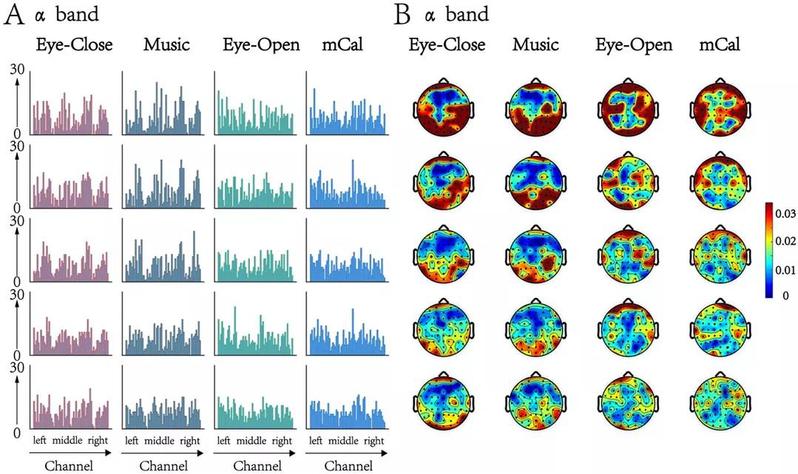
图5 | α频段中动态最小生成树核心节点分布图。(A) 所有被试动态生成树Top 5核心节点分布图;(B) 所有时间窗下所有被试的前5个最小生成树核心节点地形图分布。值得注意的是,地形图(右)中的值是左图中的频率。红色表示核心节点在20个时间窗口中出现的频率更高。
此外,动态最小生成树叶子分数 (leaf fraction, LF)、树层级 (tree hierarchy, TH) 和树直径 (diameter, D ) 结果表明这些值随时间动态变化(图6)。有趣的是,动态变异性呈现出一些稳定的趋势:LF 和D 的变异性最高出现在闭眼状态,而变异性最低则出现在心算状态。变异性衡量网络特征随时间变化的稳定性,高变异性表明大脑在这种状态下灵活变换,可随意改变信息流向呈现出联想状态;而低变异性表明大脑可能更专注于某些特定任务并过滤掉不相关信息对活跃节点的干扰。
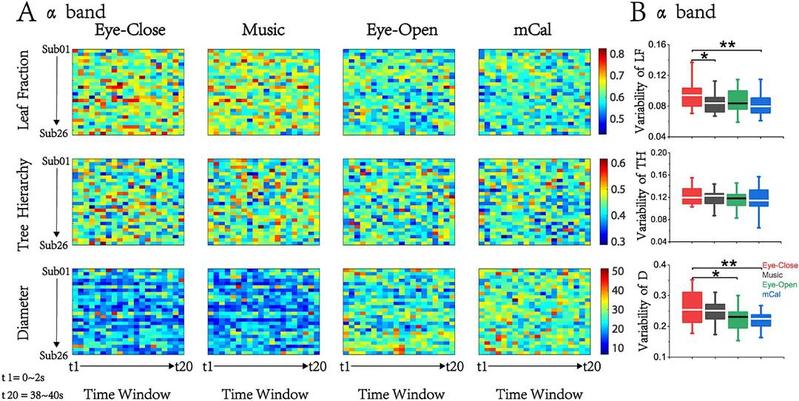
图6 | α频段中的动态最小生成树特征。(A) 动态叶子分数 (LF)、动态树层级 (TH) 和动态树直径 (D)。(B) LF、TH 和 D 的动态可变性指数。
为了进一步验证结果的可靠性,该文研究者引入了基于图拉普拉斯变换的动态不相似性指数,刻画随时间变化的最小生成树动态拓扑改变的差异性。高差异性表明大脑随着时间的推移频繁改变其状态,并且随着时间的推移,所有生成树模式之间存在非常大的差异。相比之下,低差异表明大脑随着时间的推移表现出更多的稳定性。图7结果表明,在闭眼和听音乐的状态下发现较高的差异,在睁眼和心算状态下发现动态生成树的差异较小。

图7 | 不同频段动态最小生成树的不相似性指数
通过图论动态网络特征分析,这项研究揭示人在听音乐时,大脑处于丰富多变的动态网络中,局部信息传输密集。相比之下,专注的心算任务驱使大脑进入一种更像线型的长程连接模式。在这种模式下,脑网络可实现长距离信息流通,并呈现出对局部信息流输入的屏蔽性。这些结果表明,人脑在不同认知状态下,运用了不同的网络连通模式进行高效的信息传输和计算功能。由此或可启发人工神经网络在处理不同工作任务时或可生成不同的网络连通结构来发挥联想记忆的发散特性或专注任务的高效性。将来会设计更加精巧的实验范式,以揭示更多的人脑功能网络复杂特征和规律。
Mathematical calculation usually requires sustained attention to manipulate numbers in the mind, while listening to light music has a relaxing effect on the brain. The differences in the corresponding brain functional network topologies underlying these behaviors remain rarely known. Here, we systematically examined the brain dynamics of four behaviors (resting with eyes closed and eyes open, tasks of music listening and mental calculation) using 64-channel electroencephalogram (EEG) recordings and graph theory analysis. We developed static and dynamic minimum spanning tree (MST) analysis method and demonstrated that the brain network topology under mental calculation is a more line-like structure with less tree hierarchy and leaf fraction; however, the hub regions, which are mainly located in the frontal, temporal and parietal regions, grow more stable over time. In contrast, music-listening drives the brain to exhibit a highly rich network of star structure, and the hub regions are mainly located in the posterior regions. We then adopted the dynamic dissimilarity of different MSTs over time based on the graph Laplacian and revealed low dissimilarity during mental calculation. These results suggest that the human brain functional connectivity of individuals has unique dynamic diversity and flexibility under various behaviors.
相关论文信息:https://doi.org/10.1007/s43657-021-00027-w
第一作者:复旦大学附属中山医院神经内科 郑高兴








